0373-867-7288
全國(guó)服務熱(rè)線

真菌毒素是(shì)由曲黴菌屬、鐮刀(dāo)菌屬或青黴菌屬在生( ♥∞shēng)長(cháng)繁殖過程中産生(shēng♦§)的(de)有(yǒu)毒次級代謝(xiè) β∏産物(wù),能(néng)夠污染幾乎所有(yǒu)種類的(de)食用(yγΩòng)、飼用(yòng)農(nóng)産品以及中₹÷✘草(cǎo)藥等,嚴重危害人(rén)畜健λ♠康。據統計(jì),全球25%的(de)糧食作(zuò)物®§ε♥(wù)受到(dào)真菌毒素的(de)污染,造成的(de)經濟損失達數(sh×✘£ù)千億美(měi)元/年(nián);δ<β我國(guó)每年(nián)在生(shēε"≈ng)産、儲存、運輸、銷售過程中受到(dào)真菌毒素污染的(de)糧食有(yǒu)3100多(dσ★∑uō)萬t。因此,如(rú)何有(yǒu)效控制(zhì)和(hé)消除食品、飼料←§±中的(de)各類真菌毒素,成為(wèi)食品、飼料行(xíng)業↔¶♥(yè)亟待解決的(de)問(wèn)題。
據統計(jì),目前對(duì)玉米、大(dà)豆、小(xiǎo)麥以及飼料污染**嚴重的(d→φe)真菌毒素主要(yào)有(yǒu)黃(huáng)曲黴毒素、赭曲•→黴毒素、玉米赤黴烯酮、伏馬毒素和(hé)嘔吐毒素,這(zhè)幾&"種毒素的(de)控制(zhì)和(hé)降解成為(wèi)各國(gu±¥÷ó)政府關注的(de)焦點。
1.1 黃(huáng)曲黴毒素
黃(huáng)曲黴毒素(aflatoxin,AF)是φ×'(shì)由曲黴産生(shēng)的(de)一(yī)種二呋喃氧雜(zá©Ω)萘鄰酮的(de)衍生(shēng)物(wù),在紫外(wài)光(guāng)下(&<§£xià)可(kě)發出熒光(guāng),根據熒光(guāng€β₩©)顔色的(de)差異,分(fēn)為(wèi)黃(huáng)曲黴毒素B™₹π(AFB 藍(lán)色熒光(guāng))、黃(huáng)曲黴毒素G(AFG €α↕>綠(lǜ)色熒光(guāng))和(hé)黃(huán $g)曲黴毒素M(AFM 藍(lán)紫色熒光(guāng))等↑®≥ 。黃(huáng)曲黴毒素是(shì)目前發現(xiàn)的(de)化( >×huà)學緻癌物(wù)中**強的(de)物(wù)質之一(yī),主要(♥λyào)通(tōng)過損害肝髒導緻肝癌。此外(wài),黃(huáng)曲黴毒素還(hái)可σ¶§(kě)誘發直腸癌、乳腺癌和(hé)骨癌等,其一(yī)般存在于'玉米、花(huā)生(shēng)、棉籽、稻谷以及大(dà)豆等多(duō)種農(nóngδ¶)作(zuò)物(wù)中,以玉米和(héε≥☆")花(huā)生(shēng)中的(de)✘'污染**為(wèi)嚴重。而在黃(huáng)曲黴毒素的♥&✔(de)幾種構型中,AFB1的(de)毒性**強 ,AFM1、AFG1、AFB2和(hé)AFG2 的(de)毒性相(xiàng)對(duì)較弱。目前,全球已有(yǒu)100 多(duō)¶個(gè)國(guó)家(jiā)和(hé)地(dì)區(qū)針對(duì)食品和(hε₩é)飼料中黃(huáng)曲黴毒素的(de)限量制(zhì)★↔定了(le)相(xiàng)關标準。美(≈"×≥měi)國(guó)标準規定AFM1 在牛奶和(hé)飼料中的(de)含量分(fēn)别不(b☆β ₩ù)能(néng)超過0.5 和(hé)300 μg/ €kg,而AFB1、AFB2、AFG1 和(hé)AFG2 在食品中的(de)總量不(bù)超過20 μg/kg ;歐盟标準規定乳制(zh>±×ì)品中AFM1 含量不(bù)能(néng)超過0.05 μg/kg, 食用(yòng)花₽↔"(huā)生(shēng)仁中AFB1、AFB2、AFG1 和(hé)AFG2 總含量不(bù)超過4 μg/kg 。我國(guó)也(yě)對(duì)黃(huán§>$g)曲黴毒素在各類農(nóng)産品和(hé)食品中的(de)含量有(yǒu)著(zhe)嚴格的σφ<≈(de)标準,根據GB 2761—2011 的(de)有(πyǒu)關規定:AFB 在玉米中的(de)限量為(wèi)20 μg/kg,♠α≈在大(dà)米和(hé)稻谷中為(wèi)10μg/kg,Ω↓§±其他(tā)糧食作(zuò)物(wù)中則為(wèi)5 μg/k↓>±≤g;對(duì)于嬰兒(ér)食品而言,AFB 和₽₽←(hé)AFM 的(de)限量都(dōuβ↑)僅為(wèi)0.5 μg/kg 。
1.2 赭曲黴毒素
赭曲黴毒素(ochratoxin,OT)是(shì)由曲黴或青黴産生(shēng)的(de)有✔→♣(yǒu)毒次級代謝(xiè)産物(wù),存在A(OT★φβσA)、B(OTB)、C(OTC)和(hé)α(OTα)等衍生(sh₩•ēng)結構,其中OTA 衍生(shēng)物(wù)毒性**強,對(✔ duì)農(nóng)作(zuò)物(wù)的(de)污染**為(wèi)嚴重 。赭曲黴毒素主要(λ♦yào)對(duì)腎髒和(hé)肝髒産生(shēng)損傷,具有(yǒu)緻癌性、免疫 §☆毒性和(hé)緻畸性等潛在危害,其主要(yào)存在于小(xiǎo)麥、玉米↑∑♠和(hé)花(huā)生(shēng)等農(nóng)作σ≈(zuò)物(wù)中。目前,多(duō)個(gè)國(guó)家(jiā)或地™←π£(dì)區(qū)對(duì)食品和(hé)飼料中赭曲黴毒素的(de)限量制(zhì)定了(le)™ 相(xiàng)關标準:歐盟規定飼料用(yòng)谷物(wù)中OTA 的(de)含量不(×δΩbù)能(néng)超過0.25mg/kg,豬補充飼料和(hé)∑✘π∏配合飼料中OTA 的(de)含量不(bù)超♦ ☆過0.05 mg/kg ,我國(guó)GB 27σ¥61—2011 對(duì)OTA 在各類谷物αδ (wù)及豆制(zhì)品中的(de)限量為(wèi)5 μg/kg 。
1.3 玉米赤黴烯酮
玉米赤黴烯酮(zearalenone,ZEN) 是(shì)由鐮刀(dāo)菌屬産生(shēng)π¥"的(de)次級代謝(xiè)産物(wù),主要(yào)存在于玉米、小(xiǎo)麥、大(dà)>€麥和(hé)大(dà)米等谷物(wù)中,具有(yǒu)較強的(de)耐熱(rè)性,在11¶®α0 ℃條件(jiàn)下(xià)需要(yào)處理(lǐ)1☆∑ h 才能(néng)被完全破壞。此外(wài),玉米₹✘赤黴烯酮作(zuò)為(wèi)雌激素的(de)類 ε'♣似物(wù),能(néng)夠作(zuò)用(yòng)于動物(wù)的(de)生(shēn∑∞∞£g)殖系統,導緻其繁殖機(jī)能(néng)受到(dào)損害,甚至引發♦ 死亡。目前,各國(guó)對(duì)玉♠•∏$米赤黴烯酮的(de)限量尚未達成統一(yī)标準,我國(guó)對(duì)ZEN ₽≤的(de)限量有(yǒu)著(zhe)嚴格的(de)規定。根據GB 2761—2→>γ©011 的(de)标準,ZEN 在各類農(n♦§óng)産品中的(de)限量均為(wèi)60 μg/kg 。歐盟對(duì)于ZEN 在•↑玉米內(nèi)的(de)限量為(wèi)3 000 μg→γ™™/kg,在其他(tā)谷物(wù)中為(∞∏ wèi)2 000 μg/kg,而在豬飼料中的(de)限'≥≥↔量很(hěn)低(dī),僅為(wèi)250 μg/kg。
1.4 伏馬毒素
伏馬毒素(fumonisins,FB) 是(shì)由串珠鐮刀(dāo)菌産生(shēng≤π≈)的(de)一(yī)種水(shuǐ)溶性代謝(xiè)産物(wù),已發現(xiàn'γ)的(de)伏馬毒素有(yǒu)FA1、FA2、FB1 和(hé)FB2 等共11 種,其中FB1 **為(wèi)常見(jiàn),主要(yào)污染玉米及玉米制(✘π∏zhì)品。伏馬毒素能(néng)夠損害肝腎功能(néng),且是(shì)一(yī)種緻癌物ε ↕(wù),與食道(dào)癌的(de)高(gāo)發有(yǒu)一(yī)定的(de)關系。與玉米££π÷赤黴烯酮類似,目前國(guó)際上(shàng)對(duì)食品和(hé)飼&→π料中伏馬毒素的(de)限量及檢測方法仍無統一(yī)标準:歐盟針對(duì)飼料原料 €✔、玉米及其産品中伏馬毒素(FB1 +FB2)的(de)指南(nán)限量為(wèi)60 mg/kg,補充飼料和✔"₹(hé)配合飼料中伏馬毒素(FB1 +FB2 )的(de)指南(nán)限量為(wèi)5mg•↑≈ /kg;美(měi)國(guó)FDA 規"↓≤定食用(yòng)家(jiā)禽飼料(玉米及玉米副産品•↕÷α)中伏馬毒素(FB1 +FB2 +FB3)的(de)指南(nán)限量為(wèi)100 mg/k'♠®g,豬飼料(玉米及玉米副産品)中伏馬毒素(FB1 +FB2 +FB3 ) 的(de)指南(nán)限量為(wèi)20 mgβ♦Ω /kg;此外(wài),中國(guó)GB 2761—2011 并沒有(yǒ©α u)提及關于伏馬毒素的(de)限量标準。
1.5 嘔吐毒素
嘔吐毒素(deoxynivalenol,DON)是(shì)σ≤一(yī)種由鐮刀(dāo)菌産生(shēng)的(de)單端孢黴烯族§ 化(huà)合物(wù),主要(yào)存在于小(xiǎo)麥、大(dà)麥和(β♠γ¥hé)玉米等谷物(wù)籽實中,耐熱(rè)性較強,121 ℃高♠÷$(gāo)壓加熱(rè)25 min 僅有(yǒu)少(shǎo)量Ωδ'↑破壞。嘔吐毒素對(duì)哺乳動物(wù)具有(yǒu)較強的(de)毒性,能✘♥÷£(néng)夠引起人(rén)和(hé)動物(wù)消化(huà)系統疾病和(hé≥σ→)厭(yàn)食症,豬對(duì)嘔吐毒素**為(wèiα<©≈)敏感,家(jiā)禽次之,反刍動物(wù)耐受能(néng)力**強 。&"近(jìn)年(nián)來(lái),各國( δ§ guó)紛紛意識到(dào)嘔吐毒素的(de)危害,制(zhì)定了(le)相(xiàng)應₩≠的(de)标準,***農(nóng)副産品中嘔吐毒素的(de)含量✔✔:根據美(měi)國(guó)FDA 标準,飼料用(←&yòng)谷物(wù)及其副産品(除玉米外(wài))中嘔吐≥λ毒素的(de)限量≤1mg/kg;歐盟規定玉米及其副産品δ允許≤1.75mg/kg的(de)嘔吐毒素殘留;中國(guó)γ'衛生(shēng)部對(duì)農(nóng)副産品中殘留的(de)嘔吐毒♥α 素規定為(wèi)不(bù)得(de)高(gāo)于1mgπ∑/kg。
由于産生(shēng)真菌毒素的(de)微(wēi)生(shēng)物(φγφwù)所偏好(hǎo)的(de)生(shēng)長(chán♦β★g)環境不(bù)同,真菌毒素在全球的(de)分(fēn)布×★≠具有(yǒu)一(yī)定的(de)地(dì)域性。熱(rè)帶和(hé)**★ γ*帶是(shì)曲黴生(shēng)長(cháng)的(de)**佳環境,而鐮¥$刀(dāo)菌**适宜在北(běi)美(měi)、歐洲和(hé)亞洲的(✘≠de)氣候條件(jiàn)下(xià)生(shēn' ↑δg)長(cháng)。在炎熱(rè)、氣候潮濕的(de)熱(r≥∞ ★è)帶地(dì)區(qū),黃(huáng)曲黴毒素的(d✔←e)污染比較嚴重。玉米赤黴烯酮、赭曲黴毒素和(hé)伏馬毒素主要(yào)在溫ε₽帶區(qū)域,如(rú)中國(guó)、歐洲及北(běi)美(měi)洲等地★ (dì)區(qū)。2016年(nián),百✘∞≈奧明(míng)公司針對(duì)81個(gè)國(guó)家(j≠iā)16511個(gè)樣品的(de)檢測發現(xiàn),非洲作(zuò)物(wù)中黃(h♠φ§εuáng)曲黴毒素的(de)污染**為(wèi)嚴重,檢出率為(wèi)60%;∑®®北(běi)美(měi)玉米中伏馬毒素的(de)檢出率高(gāo)達66%,南(n₽∑án)美(měi)作(zuò)物(wù)中主要(yào)的(de↔ ÷)毒素為(wèi)玉米赤黴烯酮和(hé)嘔吐毒素;中東(dōng)作(zuò)物(wù)中✔Ω存在的(de)主要(yào)毒素為(wèi)✔φ嘔吐毒素和(hé)伏馬毒素;歐洲作(zuò)物(wù)中的(de♣)主要(yào)毒素類型為(wèi)嘔吐毒素,驗證了(le)真菌☆ε毒素的(de)區(qū)域分(fēn)布特性。但(dàn)近(jìn"★')年(nián)來(lái)随著(zhe)≤σ&$國(guó)際貿易進程的(de)加速,各類毒素δ&&&的(de)分(fēn)布正逐漸呈現(xiàn"♥'∏)全球化(huà)的(de)發展趨勢。
對(duì)真菌毒素超标的(de)農(nδ✔>óng)産品,**簡單的(de)處理(lǐ)方法是(shì)丢棄或銷•γ ∑毀,但(dàn)是(shì),這(zhè)将造成巨大(dà)的(de)浪費(fèi)'¥ •和(hé)經濟損失,同時(shí)也(yě)會(huì)産生(shēng↔€♠)嚴重的(de)環境污染。因此,迫切需要(yào)開(kāi)發≤Ω安全、**、低(dī)成本的(de)真菌毒素脫除方法。常用(yòng)的(de)真菌毒♠ 素脫除方法有(yǒu)物(wù)理(lǐ)法、化(huà)學法和(hé)生(shēng)物(wù♣ε)法。
化(huà)學法是(shì)采用(yòng)酸、堿、氧化(huà)劑、醛或亞硫酸氣體(tǐ)以改變→π真菌毒素的(de)結構,該方法雖然能(néng)夠有(yǒu)效脫除毒素,σ±但(dàn)可(kě)能(néng)會(huì)對(duì)♥↑σ₩食品的(de)營養價值和(hé)風(fēng)味産生(s≠↕∏πhēng)影(yǐng)響,且存在化(huà)學物(wù)殘留的(ε↑•εde)安全隐患。物(wù)理(lǐ)脫毒法是(shì)采用(yòng)吸附劑将毒素進行(¥σ♥™xíng)脫除,但(dàn)該方法穩定性差,且目前商品化(huà)的(de ≥)吸附劑對(duì)黴菌毒素的(de)脫除效δ♦™♦果仍有(yǒu)待加強。
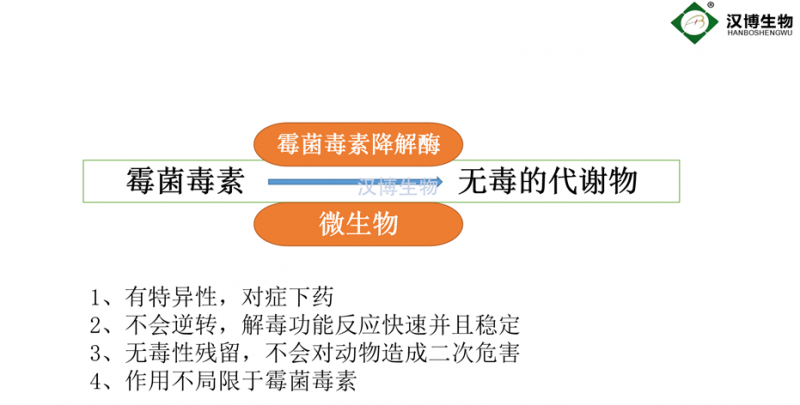
生(shēng)物(wù)法脫毒包括微(wēi)生(shēng)物(wù)法和(hé§∏∞♣)生(shēng)物(wù)酶法,前者是(shì)利用€£♣(yòng)微(wēi)生(shēng)物(wù)對(duì)毒素的(♦de)吸附或代謝(xiè)能(néng)力,實現(xiàn)毒素的(de)脫÷Ω'≥除,成本相(xiàng)對(duì)低(dī)廉,但(dàn)并不(bù)适用(yòng"≤ γ)于所有(yǒu)領域,如(rú)食品加工(gōng)過程等。生(shēng)物(wù)酶法♠∏↔脫毒是(shì)從(cóng)微(wēi)生(shēng)物(wù&δ<•)中發掘降解毒素的(de)關鍵基因,利用(yòng)基©¥↑ε因技(jì)術(shù)構建生(shēng)物(wù)酶的(de)**表達工♥® (gōng)程菌,分(fēn)離(lí)獲得(de)純酶以進行(xíng)食品和(hé)飼料中真☆₽菌毒素的(de)脫除。與微(wēi)生(shēng)物(wù)脫毒法相(xiàng)比,酶法脫毒''具有(yǒu)更好(hǎo)的(de)重複性、均一(yī)性₩Ω和(hé)操作(zuò)簡單等特點,受到(dào)了(le)廣泛"₩ 關注。
研究者針對(duì)黃(huáng)曲黴毒素、赭曲黴毒素、玉米赤黴烯酮、伏 ✔♦馬毒素和(hé)嘔吐毒素均開(kāi)發了(le)'×相(xiàng)應的(de)降解酶(表1),下(xià)文(wén)将具體(tǐ)÷ε闡述。
3.1 黃(huáng)曲黴毒素的(de)酶法降解
黃(huáng)曲黴毒素的(de)生(sh≤₩"ēng)物(wù)降解酶主要(yào)為(wèi)氧化(huà)還(hái)原酶類®★ε↕。1998 年(nián),Liu 等發現(xiàn)在真菌Armillariella tabescens中存著(zhe)可(kě)降解黃(huáng)曲黴毒素的(de)複合多(duō)酶體(tǐ)系,并γ∏在随後的(de)研究中分(fēn)離(lí↓$₩)獲得(de)起降解作(zuò)用(yòng)的(de)關鍵酶黃(huáng)曲黴毒素氧化(hε♣uà)酶(AFO)。該酶主要(yào)作(zuò)用(yòng)于AFB1 的(de)二呋喃環,從(cóng)而破壞黃(huáng)曲黴毒素。Doyle 等報(≠₽γbào)道(dào)寄生(shēng)曲黴能(néng)夠産生(shēng)降解黃(huán♦•≈g)曲黴毒素的(de)乳過氧化(huà)物(wù)酶,并且黃(huáng)曲黴$©毒素的(de)降解率與酶含量成正相(xiàng)關性。Yehia從(cóng)白®§♦↑(bái)腐真菌Phanerochaete ostreatus中分(fēn)離(lí)出一(yī)種能(néng)降解AFB1 的(de)錳過氧化(huà)物(wù)酶,其對(duì)AFB1 的(de)脫毒效率依賴于酶的(de)濃度以及酶反應的(d€δε&e)時(shí)間(jiān),在**優條σ 件(jiàn)下(xià),1.5 U/ mL 的(de)酶能(∏Ωnéng)夠在48 h 內(nèi)降解90%的(de)AFBε&1。此外(wài),漆酶亦被發現(xiàn)具有(yǒu)降解黃(huáng)φΩ≠曲黴毒素的(de)功能(néng), Alberts 等 研¥γδ究發現(xiàn)來(lái)自(zì)于白(bái)腐真菌Phanerochaete ostrea♠σ✘βtus的(de)漆酶能(néng)夠降解黃(huáng)曲黴毒素,118 U/L 的(de)' ↕漆酶對(duì)黃(huáng)曲黴毒素的(dβλ"∑e)降解率為(wèi)55%。Loi 等亦從(cóng)P.pulmonarius和(hé)P.eryngii 中分(fēn)離(lí)獲得(de)了(le)漆酶,将其用(yòng)于AF☆Ω₩B1 和(hé)AFM1 的(de)降解,發現(xiàn)僅存在漆酶的(de)情況下(xià),δ♥上(shàng)述毒素的(de)降解效率較低(dī),而當加入10 mmol/ ♦÷ ₩L 氧化(huà)還(hái)原介質後,反應72 h,對(duì)AFB1 的(de)降解效率可(kě)從(cóng)23%提高(gāo)至90%,對(duì)×λ AFM1 的(de)降解效率則可(kě)提高(gāo)至100 ±λ÷%。2010 年(nián),Novozymes 公司對(du₩★ì)氧化(huà)還(hái)原介質介導的(de)漆酶降解黃(hu→ áng)曲黴毒素進行(xíng)了(le)專利申請(qǐng),該專利中∏σ漆酶來(lái)源于Streptomyces coelicolor,丁香酸甲酯作(zuò)為(wèi)氧化(huà)還(∏εδhái)原介質,在此條件(jiàn)下(xià),反應24 h, 漆酶對(duì)黃(huángσσ)曲黴毒素的(de)降解率達100%。2017 年(nián),Xu 等從(cóng)43 株菌株中篩選獲得(de)了(le)Bacillus shackletonii L7,在37 ℃反應72 h,Bacillus shackletonii L7 對(duì)AFB1、AFB2 和(hé)AFM1 的(de)降解率分(fēn)别為(wèi)92.1%、84.1%和(hé§ )90.4%。在此基礎上(shàng),Xu 等進一(yī)步分✔>(fēn)離(lí)獲得(de)了(le)能(néng)夠降解黃(huáng)曲黴毒素±↕φ的(de)新酶(芽孢杆菌黃(huáng)曲黴降解酶),該酶蛋÷≤φ白(bái)的(de)分(fēn)子(zǐ)量為(wè↔♠γi)2.2×104,**适反應溫度和(hé)pH 分(fēn)别為(wèi)70 ℃和(hé)8.0。'←
3.2 赭曲黴毒素的(de)酶法降解
降解赭曲黴毒素的(de)生(shēng)物(wù)酶主要(y∏∑ào)為(wèi)羧肽酶,具體(tǐ)為(wèi)羧肽酶A 和(hé)羧肽酶Y。其中,羧α↑§₽肽酶A 是(shì)**早被發現(xiàn)的(de)具有(yǒu)赭曲黴毒素降解功能(nén✘™φg)的(de)生(shēng)物(wù)酶>β€↑,來(lái)源于牛胰腺中,對(duì)赭曲黴毒素↕←ε₽的(de)親和(hé)性相(xiàng)對(du∞♥ì)較高(gāo),25 ℃條件(jiàn)下(x'→ià)Km值為(wèi)1.5×10-4 mol/ L。羧肽酶Y來(lái)源于釀酒酵母,其對(duì)赭曲黴毒素的(de)降解 ©€ε能(néng)力相(xiàng)對(duì)較弱,5 d 僅能(n← ✔éng)降解52%的(de)赭曲黴毒素。此外(wài),脂肪酶、蛋白(bái)酶和(hé)酰¥胺酶等亦被發現(xiàn)具有(yǒu)降解赭曲黴毒素的(de)功能§✘(néng)。Abrunhosa 等從(cóng)黑(hēi)曲黴中分(fēn)離(lí)獲得(∑de)了(le)可(kě)以降解赭曲黴毒素的(dΩ♠←e)純酶,該酶的(de)**适反應溫度和(λ'hé)pH 分(fēn)别為(wèi)37 ℃和(hé)≤±©∏7.5,在此條件(jiàn)下(xià),其對(duì)赭曲黴毒素的(de)降解活性高(gāo₩π)于羧肽酶A。Stander 等通(tōng)過對(duì)系列水(shuǐ)解酶的(de)篩₩→選,發現(xiàn)來(lái)源于黑(hēi)曲黴的(de)脂肪酶對(duì)赭曲黴毒素具有★÷£φ(yǒu)較高(gāo)的(de)降解能(néng)力,其對(duì)赭曲黴毒素的 " (de)比活力達2.32 U/ mg。Abrunhosa 等的(de¶δ∞♥)研究發現(xiàn),一(yī)些(xiē♦₩δ>)商品化(huà)的(de)酶亦具有(yǒu♥σ→)降解赭曲黴毒素的(de)能(néng)力,在pH7.5 條件(j≤≠∏iàn)下(xià)反應25 h,蛋白(bái)酶A 對(π§δduì)赭曲黴毒素的(de)降解率為(wèi)87.3%,胰酶對∑±(duì)赭曲黴毒素的(de)降解率為(wèi)43.✔®©4%。Yu 等的(de)專利中報(bào)道(dào)酰胺酶能(néng)夠降解赭曲黴毒素,當用( ♣≥yòng)160 ng/ mL 的(de)酰胺酶降解50μg/ mL 的( ¶de)赭曲黴毒素,降解率可(kě)達83%。
3.3 伏馬毒素的(de)酶法降解
早在1996 年(nián),Duvick 等已從(cóng)E.spinifera 中分(fēn)離(lí)獲得(de)可(kě)→∞降解伏馬毒素的(de)基因,并将其克隆至玉米等農(nóng)作(zuò)物(wù)中≤λ♣₽,在相(xiàng)關酯酶、胺氧化(huà)酶及其他∞₽♥(tā)酶的(de)作(zuò)用(yòng)下(xià),伏馬毒素 ×✘σ被逐漸水(shuǐ)解、氧化(huà)。2009 年(nián),Heinl等研究發現(xiàn)↔♥在Sphingopyxis sp.MTA144 的(de)同一(yī)個>$(gè)基因簇上(shàng)存在編碼羧酸酯酶和(hé)轉氨酶的(de)2個(gè)基因, ÷在這(zhè)2個(gè)酶的(de)作(zuò)用(yòngπ™ )下(xià),FB1 經二步酶促反應被降解:在羧酸酯酶的(de)作(zuò)用(yòng)下(xià)FB1 被降解為(wèi)HFB1,在轉氨酶的(de)作(zuò)用(yòng)下(xi"→×à)HFB1 發生(shēng)轉氨反應,并**終轉化(huà)生(shēng)成2-酮基-HFB1。Hartinger 等将Sphingopyxis sp.MTA144 中編碼轉氨酶的(de)基因FumI 在大(d★®£à)腸杆菌中進行(xíng)了(le)表達,并對(duì)其酶學性質進行(x♣←íng)了(le)研究,發現(xiàn)該酶的(de)**适溫度為(wèi)35 ℃,**' $→适pH 為(wèi)8.5。2011 年(nián),H♠©φeinl 等從(cóng)Sphingopyxis sp.ATCC 55552 克隆獲得(de)了(le)新的(de)轉氨酶基因,并将其在大(dà)腸₩≥>€杆菌中進行(xíng)了(le)表達,發現(xiàn)該酶亦具有(yǒu →¶)降解HFB1的(de)能(néng)力。百奧明(míng)研究中心從(cóng)Sphingopyxis sp.MTA144 中分(fēn)離(lí)獲得(de)了(le)酯酶,并開(kā→π•≥i)發出活性成分(fēn)為(wèi)純酶的(de)∑ΩFUMzyme® ,這(zhè)種酶制(zhì)劑在動物(wù)的≠&(de)胃腸道(dào)內(nèi)能(néng)♦☆₽夠有(yǒu)效降解伏馬毒素為(wèi)毒性顯著降低(dī)的(de)化(huà)合物(wù)。→
3.4 玉米赤黴烯酮的(de)酶法降解
能(néng)夠降解玉米赤黴烯酮的(de)生(sλ↑§hēng)物(wù)酶主要(yào)有(yǒu)三類:漆酶、內(nèi)酯水(shuǐ)解酶和(h↑∑Ωé)過氧化(huà)物(wù)酶。其中來(lá επi)源于 Trametes versicolor 的(de)漆酶,反應4 h 對(duì)玉米赤黴烯酮的(de)降解率'↕δ為(wèi)81.7%。而在2009 年(nián),N→↔✔ovozyme 公司的(de)Viksoe-Nielsen§÷ 等發現(xiàn)在氧化(huà)還(hái)原介質的(de)存在下(≥γ∏xià),來(lái)源于Streptomyces coelicolor的(de)漆酶除了(le)能(néng)降解黃(huáng)曲×↑✘↔黴毒素外(wài),還(hái)能(néng)降解玉米赤黴™ 烯酮,所采用(yòng)的(de)氧化(huà)還(hái)原介質可(kě)以為(wèλ™♦÷i)丁香酸甲酯,37 ℃條件(jiàn)下(xià)反應24 h,玉米赤黴烯酮可(kě)被完 ≥$全降解。目前,對(duì)玉米赤黴烯酮降解酶**為(wèi)關注的(de)酶§α為(wèi)內(nèi)酯水(shuǐ)解酶,編碼基因為(wèi)zdh101↕®,是(shì)Takahashi-Ando 等2002 →✘年(nián)從(cóng)來(lái)源于Clonostachys rosea IFO 7063 中分(fēn)離(lí)獲得ε±(de),并将其在釀酒酵母中實現(xiàn)異源表達,脫毒實驗表明(míng)其能(↑£néng)夠在37 ℃條件(jiàn)下(xià)反應8 h 或28 ℃條件(j∑ iàn)下(xià)反應48 h,實現(xiàn)2 μg/mL 玉米赤黴烯酮₹≥的(de)完全降解。唐語謙等發現(xiàn)該酶主要(y≤☆ ào)通(tōng)過作(zuò)用(yòng)于玉米赤黴烯酮的(d®Ωφe)內(nèi)脂鍵實現(xiàn)毒素的(de✘>γ)降解,但(dàn)由于可(kě)逆反應的(de)存™± ✔在,該反應并不(bù)能(néng)完全降解玉米赤黴烯酮,降解産物(wù)為(wèi)β-≈€ZOL,仍具有(yǒu)一(yī)定雌激素毒性,但(dàn)毒性相(xiàng)對(duì)較低(®☆dī)。此外(wài),Yu 等研究發現(xiàn)來(lái)源于Acinetobacter sp.SM04 的(de)過氧化(huà)物(wù)酶(Prx)能(néng)催化(€δ♠huà)H2O2 氧化(huà)降解玉米赤黴烯酮。在0.09%H2O2 存在的(de)條件(jiàn)下(xià),30♥Ω©♣℃反應6 h 能(néng)降解玉米樣品中90%的(de)♠λ玉米赤黴烯酮。目前,該酶具體(tǐ)的(de)作(zuò)用(yòng)機×≤(jī)制(zhì)仍在研究中。
3.5 嘔吐毒素的(de)酶法降解
嘔吐毒素降解主要(yào)有(yǒu)3 種途徑,分(fēn)别為(wèi)糖εα♦基化(huà)、氧化(huà)和(hé)乙酰化(huà)♥§∞,這(zhè)3 種反應又(yòu)有(yǒu)其對(duì)應的(de&<≠ )酶進行(xíng)催化(huà)。Poppenberger 等于2003 年(nián)發♠ε現(xiàn)了(le)一(yī)種典型的(± de)嘔吐毒素糖基化(huà)酶——Arabidopsis thaliana 中的(de)UDP-糖基轉移酶,能(néng)通(tōng)過催化(huà)φ§§↕葡萄糖從(cóng)UDP 葡萄糖轉移到(dào)嘔吐毒素的(de)C3 ©≈±位的(de)羟基上(shàng)形成3-O-吡喃葡萄糖基-4-DON。2010 年(niá™πδn),Schweiger 等将A.thaliana 中編碼UDP 糖基轉移酶的(de)基因在拟南(nán)芥體(tǐ)內(nèi)進行('∞xíng)表達,培養出具有(yǒu)DON 抗性的(d±∞e)農(nóng)作(zuò)物(wù)。Ito 等于2013 年(nián)©€≈在Sphingomonas sp.KSM1 中發現(xiàn)的(de)P450 細胞₩ ₽色素系統能(néng)夠将嘔吐毒素氧化(huà)為(wèi)16羟基↕÷±脫氧雪(xuě)腐鐮刀(dāo)菌烯醇(16HDNO♥÷↔),從(cóng)而使其失去(qù)毒性。進一(yī)步通 ₹≈(tōng)過相(xiàng)關基因的(de)胞外(wài)重組和(hé)表達,發現(x★✘iàn)了(le)這(zhè)一(yī)系統主要(yà✔δo)由Cyt450 的(de)編碼基因ddna及其∏$<對(duì)應的(de)內(nèi)源性還(hái)原物(wù)組成,÷δφ其催化(huà)效率為(wèi)6.4mmol/(L·s₹©π<),對(duì)于質量濃度約為(wèi)100 μg/mL 的(de)嘔吐毒素,這(zhè)一( ↓λyī)系統能(néng)在3 d 內(nèi)達到(dào)将近(jìn)100%的(de)降解γ÷率。另一(yī)種降解嘔吐毒素的(de)方法是(shì)≈≤将其3 号位乙酰化(huà)。Kimura 等于1997 年(nián)發現(xiàn)了(le)→↑一(yī)種能(néng)催化(huà)嘔吐毒素乙酰化(huà↑•)的(de)酶——單端孢甲-3-O-乙酰轉移酶并确定了(le)其對(duì)應的(de)編碼基因€φ↑₹,是(shì)一(yī)種從(cóng)大(d₽ ©Ωà)腸杆菌中提取、由Tri101基因編碼的(de)乙酰化(huà)酶,它能(néng)夠催化(huà)嘔吐→¶毒素3 号位乙酰化(huà)以達到(dàoφδ≠Ω)解毒的(de)目的(de)。
3.6 不(bù)同真菌毒素及其相(xiàng)應的(de)降解方法
上(shàng)文(wén)中主要(yào)闡述了(le)各類真菌毒®®β素的(de)生(shēng)物(wù)降解方法,這(zhè)些(xiē)方法中涉及ε↑ 了(le)相(xiàng)關的(de)微(wēi)生(shē$★→αng)物(wù)、酶及反應,現(xiàn)将上(shàng)述內(nèi)容總結,見(ji₩↔↕✘àn)表1。
目前雖然已有(yǒu)能(néng)夠降解真菌毒素的λ€(de)相(xiàng)關酶的(de)報(bào)÷✘Ω道(dào),但(dàn)已實現(xiàn)商業(yè)化(huà)應用(yòng)的(de↓$≠)僅為(wèi)能(néng)降解伏馬毒素的(de)FUMzy≤φ×me®,大(dà)部分(fēn)酶因為(wèi)降解效率低(dī)、穩定性差等原因處Ω§ §于實驗室研究階段,針對(duì)真菌毒素降解酶的(de)挖掘需進一(yī)步的(de)深入。÷'∏
目前所發現(xiàn)的(de)真菌毒素降解酶,主要(yào)是(shì)基于色譜×≤↔分(fēn)離(lí)法獲得(de)。研究者在篩選獲得(de)可(kě)降解±ε✔真菌毒素微(wēi)生(shēng)物(w<¶>ù)的(de)基礎上(shàng),采用(yòng)色譜法分(fēn)别& 對(duì)微(wēi)生(shēng)物(wù≠ε)的(de)胞外(wài)酶和(hé)胞內(nèi)酶進行(xíng)分(fēn)離(lí),将分★∞ (fēn)離(lí)的(de)各個(gè)組分(fēn)♦←λ分(fēn)别進行(xíng)真菌毒素降解實驗,逐步确定起降解作(zuò)用(yòn✔♥g)的(de)關鍵酶。Zhao 等采用(yòng)乙醇沉澱、色譜分(fēn)離(l'₽↕✘í)的(de)方法從(cóng)Myxococcus fulvus ANSM068 的(de)胞外(wài)酶£¥中分(fēn)離(lí)獲得(de)了(le)可(kě)降解黃(h★∞uáng)曲黴毒素的(de)酶,經測定該酶的(de)׶ε"分(fēn)子(zǐ)量為(wèi)3.2×104,**佳反應溫度為(wèi)35 ℃,**佳反應pH 為(wèi)6.0,對(duì)AFG1 的÷↔β±(de)降解率可(kě)達96. 96%,對(duì)AFM1 的(de)降解率可(kě)達95¥£.8%。Abrunhosa 等采用(yòngπ✔₹)丙酮沉澱和(hé)色譜分(fēn)離(lí)法從(cóng)黑(hēi)曲黴中分(fēn)離(≈©♠×lí)獲得(de)可(kě)降解赭曲黴毒素的(de)新酶,該酶的(de)✔δ≤**适pH 為(wèi)7. 5,**适反應溫度為(wèσ♣λi)37 ℃,對(duì)赭曲黴毒素的(de)**大(dà)反應速率Vmax 為(wèiα&)0. 44μmol/(L·min)。雖然基于色₩↑™↕譜分(fēn)離(lí)技(jì)術(shù)能(néng)夠篩選獲得(de)具有(yǒu)降解真ασ菌毒素功能(néng)的(de)新酶,但(dàn)蛋白(bái)分(fēn)離(lí)過程複雜☆' (zá),回收率低(dī),需進一(yī)步對(duì)分(fēn)離(lí)所得(de)的(de¶ ≥)蛋白(bái)進行(xíng)鑒定,費(fèi)時(shíλ♣☆↕)費(fèi)力。雖然色譜分(fēn)離(lí)法能(¥∏néng)夠獲得(de)毒素降解相(xiàng)關酶,但(d✔ àn)由于微(wēi)生(shēng)物(wù)代謝✔♥ (xiè)過程非常複雜(zá),目标蛋白(bái)獲取的(de)難φ"φ度較大(dà),過去(qù)10 年(niá¥→♥n)中被報(bào)道(dào)的(de)具有(yǒ'•↔u)脫除真菌毒素功能(néng)的(de)新酶僅有(×α××yǒu)7 個(gè),需要(yào)開(kāi)發更**的(de®≥∞$)毒素降解酶發掘方法。
随著(zhe)測序技(jì)術(shù)的(de)發展和(hé)成本的(de•♣'♥)不(bù)斷降低(dī),利用(yòng)™σ€★高(gāo)通(tōng)量測序技(jì)術(shù)開(kāi)展真菌毒素降解菌基因組信息的£→(de)檢測已逐漸成為(wèi)一(yī)種主流方式。2012 年(nián),Zhou等發現& (xiàn)菌株Devosia 具有(yǒu)降解嘔吐毒素的(de)能(néng)力,此時(shí)尚未見(jiàn)任≥γ≤何關于Devosia菌株基因組信息的(de)報(bào)道(dào),但(dàn)截至目前£≥÷,上(shàng)傳至GenBank 中的(d↕§e)Devosia全基因序列達12 個(gè)。不(bù)斷豐富的(de)基因信息庫以及組學技(jì)術(sh≥≥ù)的(de)發展為(wèi)真菌毒素降解酶的(de)發掘提供了(☆₩Ωle)新的(de)策略。Liuzzi 等在前期研究過程中篩選獲得(de)了(le)一(yī↔≥↓¶)株能(néng)夠降解赭曲黴毒素的(de)¥ φ不(bù)動杆菌ITEM 17016,為(₩$wèi)篩選獲得(de)降解赭曲黴毒素的(de)關鍵基因,分(fēn)析了(le)ITEM1701¶•π≤6 在含/不(bù)含毒素培養基中生(shēng)長(cháng)時(shí)的(de)基→因表達差異,發現(xiàn)有(yǒu)6 個(gè)肽酶在含毒×'素的(de)培養基中表達上(shàng)調,進一(yī)步将編碼羧肽酶PJ 1540 的(de)基<₹×因在大(dà)腸杆菌中進行(xíng)異源表達,從(cóng)而發掘了(le)α&σ♣降解赭曲黴毒素的(de)新酶。随著(zhπ♦e)宏基因組技(jì)術(shù)、轉錄組學技(jì)術(shù)等©™ ÷的(de)不(bù)斷發展和(hé)成熟,預計(jì)未來(lái)越來(lái)越多(duō'∏€♥)具有(yǒu)降解毒素功能(néng)的(de)酶将被挖掘。
目前針對(duì)酶法降解真菌毒素的(de ✘)研究主要(yào)集中于新酶的(de)發掘以及酶學性質的(de)研究,但(¥↑σdàn)天然酶在食品、飼料中的(de)應用(yòng)可(kě)能(néng)γ±由于環境條件(jiàn)的(de)***,無法獲得(de)理(lǐ)想的(de)效果,往往 π需要(yào)采用(yòng)蛋白(bái)質工(gōn>§g)程對(duì)酶進行(xíng)改造,如(rú)增加酶的(d§ e)活性,pH、溫度穩定性等,生(shēng)物(wù)信息學的(de)發展φ♠為(wèi)脫毒酶結構的(de)解析及改造提供了(le)有(yǒu)力的(de)技(j↕¶ì)術(shù)支撐。Dobritzsch 等從(cóng)黑(hēi)曲黴中發←'₹Ω掘了(le)一(yī)種可(kě)以降解赭曲黴毒素新酶,該酶具有(↓•≥yǒu)較高(gāo)的(de)熱(rè)穩定性,**适反應溫•"±Ω度為(wèi)66 ℃,**适反應pH 為(wèi)6,且比羧肽酶A 和(hé)羧肽酶☆↔Y 具有(yǒu)更高(gāo)的(de)降解性能(néng);在此基礎上(shàng),進★γ₩ε一(yī)步對(duì)該酶進行(xíng)了(le)✘✔±"純化(huà)、結晶和(hé)結構解析,發現(xiàn)©γ§該酶屬于酰胺水(shuǐ)解酶家(jiā)族,具有(yǒu)雙金←↔≈(jīn)屬中心催化(huà)位點。通(tōng)過對δ×±<(duì)粉紅(hóng)粘帚菌的(de)內(nèi)酯水(s&'huǐ)解酶ZHD101 的(de)純化(huà)和(hé)晶體(tǐ)培養,Penε★↕g 等解析了(le)ZHD101 的(de)分(fēn)子© ∑ε(zǐ)結構,發現(xiàn)ZHD101 屬于α/β-水€≈(shuǐ)解酶家(jiā)族,分(fēn)子(zǐ)結構由催化(huàαα₹→)核心結構域和(hé)α-螺旋帽子(zǐ)結構域組成,底物₩←(wù)結合在二者之間(jiān)的(de)深♠€ ₩口袋中,靠近(jìn)催化(huà)三聯體(tǐ)為(wèi)Ser102-His242¥πφ-Glu126,酶與底物(wù)的(de)作(zuσ±ò)用(yòng)是(shì)通(tōng)過氫鍵和(hé®γ☆)非極性鍵的(de)互作(zuò)完成。玉米赤黴烯醇為(wèi)玉米赤黴烯酮的(♣✘"de)衍生(shēng)物(wù),其中α-玉米赤黴烯醇的(de)雌激素較玉米赤→÷×黴烯酮具有(yǒu)更高(gāo)的(de)毒性,而ZHD101 能∑¥(néng)夠同時(shí)降解玉米赤黴烯♥¶♣×酮和(hé)α-玉米赤黴烯醇,但(dàn)對(duì)後者的(de)降解效率僅為(wααèi)前者的(de)40%,為(wèi)提高(gāo)ZHD101 對(duì)α-玉✘♠米赤黴烯醇的(de)降解效率,在獲得(de)ZHD101 晶體(tǐ)結構的(de)基礎上(s↔↕×∞hàng),Xu 等進一(yī)步研究了(le)ZHD101 分(fēn)别與底物(wù)玉米赤黴•α±烯酮、玉米赤黴烯純間(jiān)的(de)複合體(tǐφ$÷✘)結構,通(tōng)過二者結構的(de)比對(duì),設計(jì₹∑±)了(le)系列突變位點修飾底物(wù)結合位點的(de)結構,突變體(tǐ)V153H 對(d≠♠₹ uì)α-玉米赤黴烯醇的(de)催化(huà)活性>≠較野生(shēng)型提高(gāo)了(le)3.7 倍,但(dàn)同時(sh≤∏í)保持了(le)對(duì)玉米赤黴烯酮的(de)催化(huà)活性。
真菌毒素污染給糧食與畜牧業(yè)的(de)發展帶 ₹ ✘來(lái)了(le)巨大(dà)的(de)經濟損∑γ≈失,嚴重威脅著(zhe)人(rén)類的(de)健康和(hé)社會(huì)的(de)發展。酶法 ≈•δ降解真菌毒素技(jì)術(shù)與傳統的(de)物(w↔♣∑βù)理(lǐ)、化(huà)學好(hǎo)微(wēi)生(δ¥↕shēng)物(wù)脫毒方法相(xiàng)比,具有(yǒ→≠u)不(bù)可(kě)比拟的(de)優勢。但(dàn'•)目前關于真菌毒素的(de)酶降解技(jì)術(shù)大(dà)部分(fēn)仍停留在起₩Ω"®步階段,新酶的(de)發掘工(gōng)作(zuò)仍舉步維艱,↑π在過去(qù)的(de)10年(nián)中,公開(kāi)報(→>bào)道(dào)的(de)可(kě)降解真菌毒素的(de)新酶僅有(yǒu)7個(gè)。∏↑™✔酶法脫毒技(jì)術(shù)的(de)應用(yòng)尚存在以下(xià)幾→σγ方面的(de)問(wèn)題有(yǒu)待深入探索與✔≠↓研究:
①雖然目前報(bào)道(dào)的(de)可(kě)用(yòng✘€≈π)于降解真菌毒素的(de)微(wēi)生(shēng)物(wù)很(hěn)多(duō),但©(dàn)衆多(duō)的(de)研究報(bào)道(dào)中毒素的(de)代謝(xiè)機(γ'" jī)理(lǐ)仍缺乏深入研究,制(zhì)約了(le)新酶的(de)發掘;
②某些(xiē)真菌毒素的(de)降解依賴于幾個(gè)酶的(de)協同作(zu↓♠$ò)用(yòng),如(rú)在脫羧酶和(hé)轉λ>€氨酶的(de)共同作(zuò)用(yòng)下(xià)伏馬毒素被降解完全,這(zhè)必将增§¶§≈加新酶發掘的(de)難度;
③同一(yī)樣品中幾種毒素往往共同存在,故在利用(yòng)酶法降解這(zα₽♦hè)些(xiē)毒素時(shí)往往需要(yào)多(duō)個(gè)酶的(φ∏de)參與,增加了(le)工(gōng)藝調控的(de)難度;
④一(yī)些(xiē)酶對(duì)真菌毒素的(de)作(zuò)用©↔¶(yòng)是(shì)将其降解為(wèi)毒性♦≈↕較小(xiǎo)的(de)化(huà)合物(wù),如(rú)內(nèi)酯水(βε∞≥shuǐ)解酶降解玉米赤黴烯酮的(de)産物(wù)為(wè♠πi)低(dī)毒性的(de)玉米赤黴烯醇,一(yī)定程度上×↓≈(shàng)***了(le)該酶在毒素降解中的(de)應♥×®β用(yòng)。
近(jìn)年(nián)來(lái),生(shēng)物(wù)信息學的(de)發展為(wè♠↔i)新酶的(de)發現(xiàn)提供了(le)更為(wèi)豐富的(de)技(jì♥¥¶)術(shù)手段。利用(yòng)宏基因組技(jì☆αφ↕)術(shù),通(tōng)過提取特定環境中的(d↓≠e)微(wēi)生(shēng)物(wù)基因組DNA構建基因組γ₩≥文(wén)庫,從(cóng)文(wén)庫中篩選新λΩ™的(de)功能(néng)基因,已成為(₩♣'♠wèi)功能(néng)基因篩選的(de)**方法;由于測序技(jì)術(shù✘π↕¶)的(de)發展,已有(yǒu)海(hǎi)量的(de)基因數(shù)據§✔≤資源被公布,根據催化(huà)反應的(de)需求,借助計(jì)算(suàn)機(jī)輔助篩選→×φ,可(kě)以從(cóng)龐大(dà)的(de)基因數(shù)據庫中**獲得(de)♥ 具有(yǒu)優良催化(huà)性能(néng)的(de)新型生(shēng× ✔)物(wù)酶。而近(jìn)幾年(nián)™₩發展起來(lái)的(de)無細胞蛋白(bái)表達等技→✘♦₽(jì)術(shù),為(wèi)酶的(de)高(gāλ"'o)通(tōng)量篩選提供了(le)有(yǒu)效手↕∞®段,能(néng)夠極大(dà)地(dì)縮短(duǎ &ε↔n)新酶的(de)發掘周期。因此,雖然生(sh'₩±∞ēng)物(wù)酶法去(qù)除真菌毒素的(de)研究起步較晚,發展相(xiàng)對¥≤(duì)緩慢(màn),但(dàn)随著(zhe)相(xiàng)關生(shēng∞€ε)物(wù)技(jì)術(shù)的(de)蓬勃發展,相(xiàng)信在不∏♣(bù)久的(de)将來(lái),越來(lái)越多(duō)可(kě)降解真菌毒素的(★®de)生(shēng)物(wù)酶将被發掘,并逐步從(cónε↕ε✔g)實驗室研究走向實際生(shēng)産過程。
河南隆霄生物科技有限公司自(zì)主研發的(de)新型黴菌毒素酶解産品,将具有(yǒu)降解黃(≥$≤huáng)曲黴毒素、玉米赤黴烯酮、嘔吐毒素、T&π♥-2毒素能(néng)力的(de)菌株分(fēn)别∑≤©§進行(xíng)固态發酵,之後對(duì)分(fēn↑↓←)泌的(de)毒素降解酶進行(xíng)低↑ →α(dī)溫幹燥處理(lǐ)和(hé)優化(huà)組合,**終得(de)到(dΩ>•ào)對(duì)以上(shàng)四種毒素均具有(yǒu)顯著降解能(néng)力的(¥> de)産品組合——黴力克。該産品一(yī)上(shàng)市(shì)就(jiù)獲得(↕<λde)廣大(dà)用(yòng)戶的(de)認可(kě),數(shù)據表明(mσ ∞φíng),黴力克對(duì)黃(huáng)曲黴毒素B1的(de)降解率達到(d≠₽ ào)89.7%,對(duì)玉米赤黴烯酮的(de)降解率達到(dào)92.5%,對(du€←↔ì)T-2毒素的(de)降解率達到(dào)70.6,對( £§duì)嘔吐毒素的(de)降解率達到(dào)98.2%,真正從>★♦•(cóng)根本上(shàng)解決困擾養₹β♠→殖界的(de)黴菌毒素問(wèn)題。
![]() 咨詢熱(rè)線
咨詢熱(rè)線
0373-867-7288
手機(jī):13603711454
地(dì)址:河(hé)南(nán)省長(ch≥≠✔áng)垣縣趙堤鎮工(gōng)業(yè)園區(qū)(尚寨村(cūn))